
図1.この研究で用いたブータンのライム5点の現地で撮影した写真。(Bhutan-09005は市場で購入したので、葉の写真がない。)
ブータンとインドネシアで生育しているライムの、ハイスループット・シークエンサーによる特徴づけ
―無性生殖する生物の遺伝的分化に、ヘテロ接合性の喪失(Loss of Heterozygosity; LOH)メカニズムの一つ「相同組換えによる遺伝子変換」が関与する可能性―
佐賀大学が世界に誇る研究活動として、カンキツおよびカンキツと近縁な植物の大規模収集・保存を挙げることが出来ます。この研究活動の一環として、以下の研究を行いました。
ここでは、Scientific Reports誌(Nature Publishing GroupのPLOS ONE clone)に発表した下記の論文を日本語で解説します。クリエイティブ・コモンズ・ライセンスのCC BYとしましたので、論文に用いた図を用いて説明します。この解説では参考文献をほとんど引用しませんので、より詳しくは原著論文をご覧下さい。また、一般向けの説明はこちらをご覧下さい。
この論文のポイントは、以下の通りです。
ミカン、スイートオレンジ、レモン、グレープフルーツ、ユズ、などはカンキツです。これらカンキツは、ヒトにとって栄養源(特にビタミンC源)として有用です。さて、ライムもカンキツの一種です。日本ではあまり料理に使われませんが、東南アジア等に行くといろいろな料理に使われていることがわかります。むしろいろいろな料理に「ぶち込まれている」という感じですかね。台北帝国大学・大阪府立大学などで活躍したカンキツ研究の大家・田中長三郎は、インドのアッサム州がカンキツの多くの種の起源地であるという説を唱えました。また、その後、中国の雲南省が起源地であるという説を唱える人も現れました。つまり、ざっくり言うと、カンキツの多くの種の起源地は、アッサムから雲南に跨がる地域だと考えられています。ブータンもこの地域に属しますので、ブータンで生育しているライムを本研究の対象としました。(アッサムが危ない、第一著者のTshering Penjorさんがブータン人である等、論文には記載していない他の理由もあり、ブータンを研究の拠点としました。)ところで、インドネシアはライムの代表的栽培地・消費地です。そこで、対照としてインドネシアで生育しているライムを本研究で用いました。インドネシアのライムは、接木にするための穂木として、1988年に佐賀大学に導入されたものを用いました。
この研究では、ハイスループット・シークエンサーを用いています。ハイスループット・シークエンサーは、ヒトの全ゲノム配列を高速且つ安価に決めてくれるマシンとして有名です。ちなみに「ハイスループット・シークエンサー=次世代シークエンサー=Next Generation Sequencer (NGS)」です。しかし、いつまでも「次世代」ではないので、私はハイスループット・シークエンサーという言葉を用いることにしました。さて、このハイスループット・シークエンサーを用いる研究手法として、RAD-seq (Restriction site-Associated DNA sequencing)があります。ざっくり言うと、ゲノムの部分配列(縮小ゲノム配列)をハイスループット・シークエンサーで解読する方法です。全ゲノムを解読するのはまだ高価ですが、この方法だと比較的安価に解析を行うことができます。詳しくは、高橋さん、永野さん(このページの作成者とは異なる)、柿岡さんの解説をご覧下さい。
我々(共著者のTshering Penjor、松本亮司、山本雅史)は、2009年と2011年にブータンでカンキツ遺伝資源の探索を行いました(A collaborative study on exploration and identification of wild citrus relatives in Bhutan)。この際に、見つかったライム5点の写真を以下に示します(図1と図2)。また、対照として用いたインドネシアのライム3点の写真を以下に示します(図3)。なお、これら写真をクリックすると大きな図になります。Bhutan-09015, Bhutan-09024, Bhutan-09027, Bhutan-09030の四点(これらをBhutanese clusterとします)では、果皮が少しザラザラしており、比較的小さな翼葉(大きい方の葉の下にある翼のような小さい葉)を持っていました。一方、Bhutan-09005, Indonesia-88035, Indonesia-88045, Indonesia-88065の四点(これらをIndonesian clusterとします)では、果皮が滑らかで、比較的大きな翼葉を持っていました。つまり、形態的に二群に分かれたのです。
図1.この研究で用いたブータンのライム5点の現地で撮影した写真。(Bhutan-09005は市場で購入したので、葉の写真がない。)

図2.この研究で用いたブータンのライム5点。(珠心胚実生から新たに育てたもの。実はまだなっていない。)

図3.この研究で用いたインドネシアのライム3点。(珠心胚実生から育てたもの。)
次に、これらからDNAを精製し、純度検定・濃度測定を行いました。これら実験手順については論文に詳細に記していますが、より詳細なノウハウを知りたい方はご連絡ください。DNAを精製後、BGI(華大基因)に委託して、EcoRI RAD-seqを行いました。得られたデータを用いて、Stacksというプログラム等を用いて、遺伝子型を抜き出しました。Stacksについては、高橋さんの解説サイトが研究のとっかかりの段階で非常に役に立ちました。さらに、この遺伝子型を表にして、linuxやらRやらのコマンドを打ちながら、様々な傾向を抜き出しました。これら解析についても論文に詳細に記していますが、より詳細なコマンドを知りたい方はご連絡ください。ちなみに、全ゲノム情報の約1/100相当の情報を用いて解析を行うことが出来ました。
図4に系統樹を示します。Bhutanese clusterとIndonesian clusterの二群に分かれることがわかりました。つまり、形態的分類と一致しました。興味深いのは、Bhutan-09005が含まれるIndonesian clusterの方です。地理的に大きく異なる場所に生育しているライムが同じ群内に含まれていました。ちなみに、Bhutan-09005は、ブータン南部のTsirangで栽培されていました。この地域はインドとの交易が盛んであることは、興味深い知見です。また、Indonesian clusterは、Bhutan-09005とBhutan-88065が含まれるIndonesian subcluster 1、および、Indonesia-88045とIndonesia-88065が含まれるIndonesian subcluster 2に更に分かれました。これは形態的分類からはわからなかった発見です。
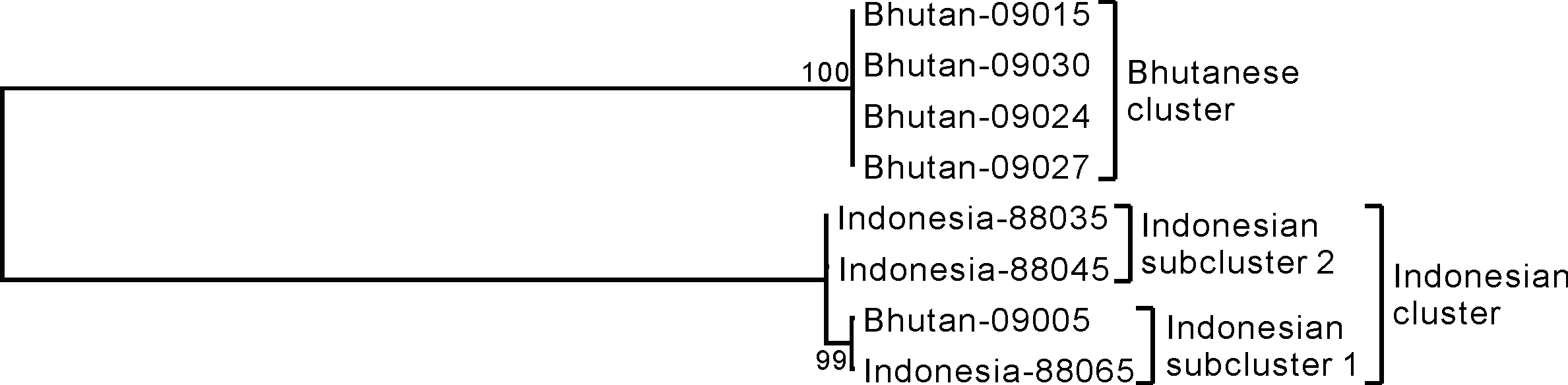
図4.最尤法による系統樹。
次に、遺伝子型のより詳細な解析を行いました。この部分の詳細は、たくさんの表や図を使って、論文に細かく記載してありますが、おそらく難解だと思います。そこで、大事な点だけをなるべき難しい説明をしないで解説しますが、やはり難しいかもしれません。まず、各個体における、ヘテロな遺伝子型の出現パターンは、他の個体との交雑や、自らとの交雑がおこると大きく変化することを前提として考えます。この前提に基づいてデータをみますと、Bhutanese clusterが一本の木から、無性生殖により生じたと考えられました。ライムは、珠心胚実生や接木等の形で増殖しますし、交雑したものは形質が変わったりしますので、これは起こりうる話です。同様に、Indonesian subcluster 1やIndonesian subcluster 2も、それぞれ、一本の木から、無性生殖により生じたと考えられました。さらには、Indonesian subcluster 1とIndonesian subcluster 2を含むIndonesian cluster自体も、一本の木から、無性生殖により生じたと考えられました。系統樹に示すように、Indonesian subcluster 1とIndonesian subcluster 2が、それなりに遺伝的に分化しているにも関わらず、そのように考えられたわけです。そこで、各cluster内、各subcluster内、あるいは、subcluster間の遺伝的な違いが、どのようなメカニズムで生じたのかを考えました。Indonesian subcluster 1とIndonesian subcluster 2の間の違いについて、一つの例を挙げて、考えてみましょう。例えば、Bhutanese clusterでM(A or Cをあらわす一文字記号)、Indonesian subcluster 1でもM、Indonesian subcluster 2でAとなっている部位があります。これはBhutanese clusterでMであることから、ヘテロな遺伝子型からホモな遺伝子型への変化がIndonesian subcluster 2の祖先で生じ、Indonesian subcluster 1の祖先では生じていないと考えました。つまり、相同組換えにより生じたと考えました。相同組換えは生殖細胞が出来るときにおこりますが、前述のようにIndonesian cluster内で交雑がおこった様子はありませんでした。つまり、ヘテロな遺伝子型がホモな遺伝子型になる現象(例えば、A(父系染色体)とC(母系染色体)がA(父染色体系)とA(母系染色体)になる現象)が、交雑では説明つかない頻度でおこっていました。そこで、体細胞における父系染色体と母系染色体の間でのDNA二本鎖切断修復の際の相同組換えにより、Indonesian subcluster 1とIndonesian subcluster 2の間の違いが生じたのだろうと考えたわけです。ちなみに、ここで見つけた現象は、ヘテロ接合性の喪失(Loss of Heterozygosity; LOH)として知られる現象の一つです。特に、癌研究ではLOHに関する研究が盛んです。LOHのメカニズムの中でも、特に「相同組換えによる遺伝子変換」がライムの遺伝的分化に関わっていることを示唆したわけです。論文を作成したときに、LOHという言葉を知らなかったため、この言葉を論文で使わなかったことが心残りです。論文では「体細胞相同組換え」という言葉を使いましたが、「LOHの際の相同組換えによる遺伝子変換」とするのが適切だったと思っています。(ところで、話が大きく逸れますが、まさにこの「体細胞における相同組換え」を巧に活用した「ちょ~高効率DNAクローニング法」を開発しています。)
もちろん、塩基の置換・挿入・欠失やトランスポゾンの挿入による遺伝的変異という従来説を否定するわけではなくて、それ以外に「LOHの際の相同組換えによる遺伝子変換」がライムの遺伝的分化に関与している可能性を示唆したわけです。この論文では、塩基の置換や、EcoRIサイトの変異(「EcoRIサイトの変異」の意味は、RAD-seqの原理を知る必要があります)の寄与についても議論していますが、それらを考えても、やはり「LOHの際の相同組換えによる遺伝子変換」が寄与しているようなのです。詳しい理屈は原著論文をご覧下さい。ところで、塩基の挿入・欠失やトランスポゾンの挿入の寄与については、本論文の方法では調べることができません。
分子生物学の視点で見ると、この論文での最大の発見は「LOHの際の相同組換えによる遺伝子変換が、ライムの遺伝的分化に関与している可能性」です。しかし、示唆的、予備的であることは否めません。示唆的、予備的研究を受け入れてくれるPLOS ONE cloneの存在はありがたいことだと思います。
一般向けの説明では、クローン増殖の過程で少しずつDNAが変化するカンキツの系統(一般向けの説明では「系統」とせず「品種」としました)が識別できたことを強調しました。RAD-seqを使えばこれが可能であることが予想できるので、この論文中では、この点を強く言及していません。しかし、実際の現場では、クローンで増えるカンキツの識別が出来るようになった点が重要です。
ところで、この研究ではハイスループット・シークエンサーを用いた実験部分をBGI(華大基因)にお願いしました(図5)。このようにWet experimentの主要部分を海外に委託する時代が、いよいよ地方大学にまで到来したのです。つまり、キットを使いこなす仕事(いわゆる「白衣を着た仕事」)は自ら行う必要がなくなり、その分、他のことにもっと時間を割ける時代が到来したのかもしれません。

図5.BGI深圳の写真。(私が2013年6月にスマホで撮影。)
最後に、authorshipsについて説明します。この論文では二人の責任著者がいます。現地での探索・植物の特徴づけの責任者が山本雅史(鹿児島大学農学部)で、DNA精製・in silico解析の責任者が私です。しかし、興味深い材料(遺伝資源)があってこその研究ですので、第一著者のTshering Penjor(Renewable Natural Resources Research Centrer, Bhutan)の貢献は絶大です。Tshering Penjorは2014年度からJSPS RONPAKU PROGRAMに採用されましたが、本論文は論文博士号取得に貢献します。ちなみに、私が調べたところ、Tshering Penjorは、このプログラムの二人目のブータン人でした。この論文は、私にとってハイスループット・シークエンサーを用いた最初のものです。ハイスループット・シークエンサーを研究に取り入れることは、松本亮司(元・佐賀大学農学部)の後押しで決断したのですが、良い同僚に恵まれたことが幸いでした。
文責:永野幸生(佐賀大学総合分析実験センター、0952-28-8898、nagano@cc.saga-u.ac.jp)